Peto's Paradox
 Whales, Elephants, and Bats
Whales, Elephants, and Bats
One of the many observations we have made is frequently referred to as Peto’s Paradox, so named after Richard Peto, who showed in 1975 that, per cell, mice had a higher rate of cancer than humans, and mused about the relationship between body-size and cancer rates. The Paradox, in a nutshell, comes from the simple fact that every cell in an animal’s body has the potential to become a tumor given the right set of mutations; furthermore, cells will steadily accumulate mutations throughout their lifetime. Thus, if you compare two species - large or small, long-lived or short-lived - you would expect the one with more cells, and a longer lifespan, to have an astronomically larger incidence of cancer. However, the fact that whales, elephants, and other long-lived and/or enormous animals have cancer rates comparable or below our own as humans, means that there must be some other mechanisms at play which compensate for the increased cancer risk; somehow, these species are able to either lower their own per-cell cancer rates, or else detect and destroy tumors near their onset.
This project was completed during my graduate career at the University of Chicago, and was pursued under the tutlage of Vincent J. Lynch, who is now at the University of Buffalo. My thesis, titled “The Role of Gene Duplication in Mediating Peto’s Paradox in Afrotheria and Chiroptera,” explores this question in great depth using a functional genomics and comparative biology approach that highlights the diversity of skills I developed in the Lynch Lab. What follows is the abstract to my thesis.
Cancer is a disease intrinsic to multicellularity. Within a species, body size and cancer risk are strongly correlated with cancer risk; between species, however, this correlation no longer holds. This phenomena, known as Peto’s Paradox, requires that species evolve cancer suppression mechanisms alongside increases in size and lifespan. Previous candidate gene studies looking at tumor suppressor duplications in large, long-lived species point towards a greater role of gene duplication in resolving Peto’s Paradox. Thus, in this thesis, I characterized gene duplications genome-wide in publicly-available genomes to determine if tumor suppressor genes in particular were enriched among duplicated genes in large and/or long-lived species. Then, to explore the functional consequences of these duplicates, I selected two hits in large, long-lived species - LIF in the African Savanna Elephant (Loxodonta africana) and TP53 in the Little Brown Bat (Myotis lucifugus) to characterize in primary fibroblasts, and determine their effects on cell cycle and cell death in response to stress.
Expanding upon previous candidate-gene-based surveys of gene duplication in Elephants, we used a Reciprocal Best-Hit BLAT strategy to obtain copy numbers of all protein-coding genes in Atlantogenatan genomes to see if there is any correlation between the copy number of duplicates and changes in body size along the phylogenetic tree. From an initial set of 18,011 protein-coding genes in hg38, we identified a median of 13,880 genes in Atlantogenatan genomes, of which a median of 940 genes are duplicated. We find that, just as body size fluctuates throughout Atlantogenata, genes involved in tumor suppressor pathways are also duplicated throughout the phylogenetic tree. Extant species of elephants, however, show active transcription of both canonical and duplicated copies of tummor suppressors that duplicated prior to and during their sudden increase in body size, suggesting that the duplication of tumor suppressor genes facilitates the evolution of increased body size by compensating for the increased cancer risk.
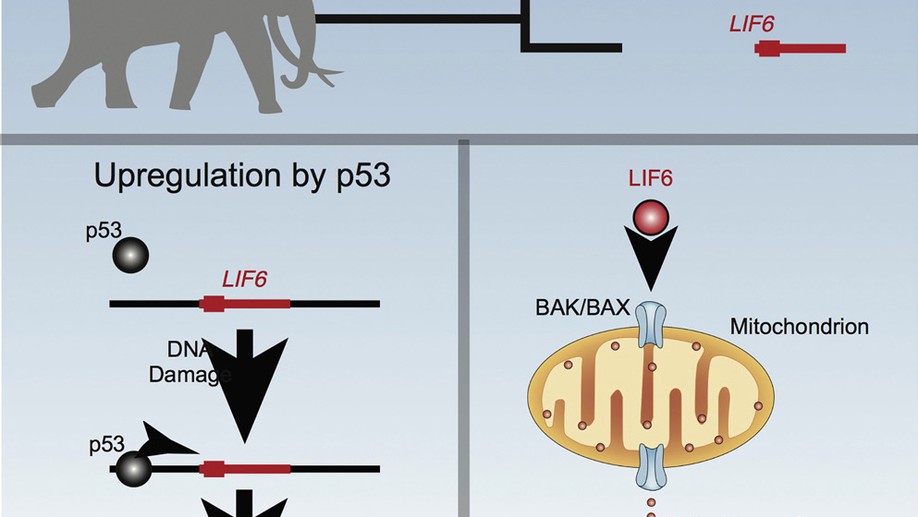
The resurrection and re-functionalization of a leukemia inhibitory factor pseudogene (LIF6) with pro-apoptotic functions in elephants and their extinct relatives (Proboscideans) may have played a role in resolving Peto’s Paradox. LIF6 is transcriptionally up-regulated by TP53 in response to DNA damage, and translocates to the mitochondria where it induces apoptosis. Phylogenetic analyses of living and extinct Proboscidean LIF6 genes indicates its TP53 response element evolved coincident with the evolution of large body sizes in the Proboscidean stem-lineage. These results suggest that re-functionalizing of a pro-apoptotic LIF pseudogene may have been permissive (though not sufficient) for the evolution of large body sizes in Proboscideans.
In the long-lived bat, Myotis lucifugus, I describe the only known full-locus duplication of TP53 in nature, which may play a role in shaping its unique stress response. This bat has 8 copies of TP53, a central regulator of the DNA damage response present in all living organisms. A syntenic duplication of TP53-WRAP53 has occurred within its genome, leading to two copies of TP53 that have conserved both regulatory and transcriptional functionality. Relative to 4 other closely related bat species (M. evotis, M. thysanodes, M. yumanensis, and E. fuscus), the Little brown bat demonstrates a unique resistance to both DNA damage as well as generalized oxidative stress, which matches the phenotypic changes induced by a transgenic TP53 locus duplication in a previously-described mouse model.
Overall, these results suggest that gene duplication plays an important role in Peto’s Paradox; while individual cases of tumor suppression duplication can facilitate the evolution of increased lifespans and body sizes, the suppression of cancer risk likely requires a concert of genetic changes. This fits in with a polygenic or omnigenic model of cancer resistance and tumorigenesis, and suggests that large-scale integrated functional genomic approaches to studying Peto’s Paradox may provide greater clarity into how large, long-lived species suppress their increased cancer risk.
Juan Manuel Vazquez
Postdoctoral Researcher
A recent graduate of the University of Chicago, I study the evolution of lifespan and cancer resistance in exceptional animals such as bats, elephants, and whales.
Publications
A Full-Locus Duplication of TP53 Enhances the Stress Response of the Little Brown Bat, *Myotis lucifugus*
Bats are extraordinarily long-lived relative to other mammals; among them, Myotis lucifugus is one of the longest-lived bats. While M. lucifugus and other closely-related species of bats have been the focus of research for a long time, there have been no reported cases of cancer in the literature - despite their long lifespan - suggesting that they have a low lifetime risk of cancer. In this bat, we find that there is a duplication of the TP53-WRAP53 locus, which is the only such known duplication among sequenced species. The two loci show active transcription both in vivo and in vitro in primary fibroblasts, suggesting that they are both functional. The responses to DNA damage response in M. lucifugus relative to its closest relatives is reminiscient of the effects of a TP53-WRAP53 duplication in transgenic mouse models, suggesting that this duplication may play an important role in mediating the cancer resistance of M. lucifugus.
Pervasive duplication of tumor suppressor genes preceded parallel evolution of large bodied Atlantogenatans
While body size and lifespan directly impact an individual’s cancer risk within species, we see no such correlation when comparing cancer risk between species - a surprise that is known as Peto’s Paradox. While there are many ways that Evolution can resolve this paradox, gene duplication stands out as a particularly parsimonious solution to the problem. Inspired by previous test cases where a tumor suppressor gene duplication was found in a large, long-lived species - such as in elephants - we sought to test whether or not tumor suppressor duplicates are especially enriched among duplicated genes in large Atlantogenatans. We find that tumor suppressor duplicates are present in all Atlantogenatan genomes, and occured throughout the tree. Tumor suppressor duplicates in Elephants show functional transcription, suggesting that these duplicates have preserved a functional role, and may have permitted the sudden increases in body size we observe throughout Atlantogenata.
The Role of Gene Duplication in Mediating Peto's Paradox in Afrotheria and Chiroptera
Based on emperical studies of humans, mice, and various other species, an individual’s cancer risk is directly proportional to their cell count (body size) and lifespan. This leads to a theoretical prediction that large and/or long-lived species would possess a higher predisposition to cancer compared to smaller, shorter-lived species; compounding this risk is the fact that body size and lifespan are strongly correlated. However, in a phenomenon known as Peto’s Paradox, cancer risk between species does not correlate with either their body sizes or lifespans. This implies that enhanced cancer resistance mechanisms must co-evolve with increases in body size and lifespan; however, there are many ways this can come about. Rather than reinventing the wheel, species can carry an increased load of cancer risk by increasing the number of wheels they have. My thesis focuses on the role tumor suppressor gene duplications play in Peto’s Paradox: Chapter 1 explores whether or not tumor suppressor genes are especially enriched among duplicated genes in large, long lived species, while Chapters 2 and 3 functionally characterize two such duplications. Overall, my work here highlights the vital role that tumor suppressor gene duplicates play in lowering the cancer risk of large, long-lived species, while also highlighting new questions for future work, especially regarding antagonistic pleitropy and growth-suppression paradoxes with these duplicates.
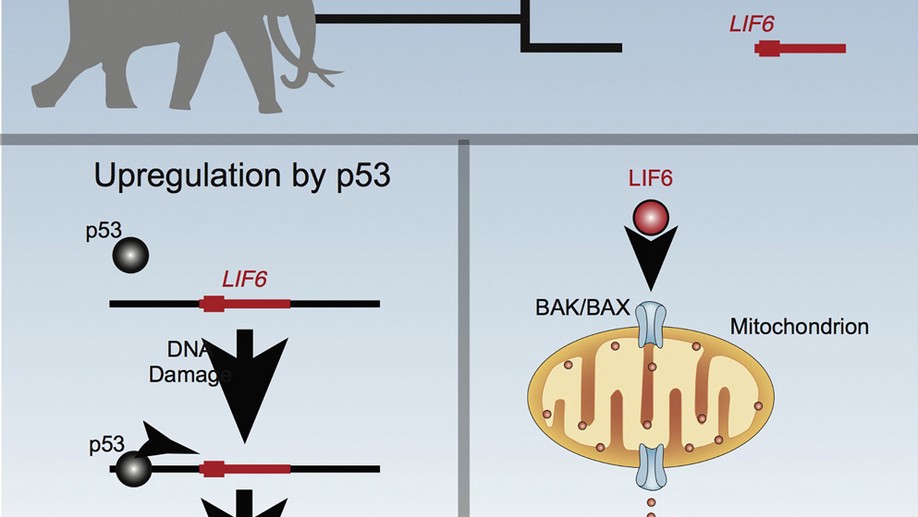
A Zombie LIF Gene in Elephants Is Upregulated by TP53 to Induce Apoptosis in Response to DNA Damage
Larger organisms with increased cell counts have a theoretically increased risk of cancer; the observation that larger species do not seem to have an increased cancer risk in contradiction to the patterns observed between members within species is known as Peto’s Paradox. ere, we show that elephants and their extinct relatives (proboscideans) may have resolved Peto’s paradox in part through refunctionalizing a leukemia inhibitory factor pseudogene (LIF6) with pro-apoptotic functions. LIF6 is transcriptionally upregulated by TP53 in response to DNA damage and translocates to the mitochondria where it induces apoptosis.